
Zurück in die Zukunft
Die Gene lassen in die Vergangenheit der Pflanzen blicken und verraten ihre künftigen Entwicklungswege
von Marcus Koch
Es gibt kaum noch Stimmen, die den aktuellen Klimawandel leugnen. Ebenfalls kaum noch zu ignorieren ist, dass mit dem Klimawandel ein weltweites Artensterben einhergeht, von dem sehr viele Organismen betroffen sind. Wie sich dieses vor unser aller Augen stattfindende Sterben auf die Ökosysteme der Erde auswirkt, wird hingegen kontrovers diskutiert. Besonders schwierig ist es, künftige Entwicklungen vorherzusehen. Unsere Forschungsarbeiten zeigen, dass ein Blick zurück in die Vergangenheit gut geeignet ist, um künftige Entwicklungen vorauszusagen. Dabei haben wir überraschende Zusammenhänge aufgedeckt, und es ist zu hoffen, dass aus dem neuen und tieferen Verständnis vom Entstehen, Werden und Vergehen der biologischen Vielfalt auf Erden nachhaltige globale Strategien zum Schutz des Lebens hervorgehen werden.
Den „Blick zurück in die Zukunft“ erlauben neue Techniken, die der molekularbiologischen Revolution zu danken sind. Sie hat vor kaum mehr als 20 Jahren weite Teile der Naturwissenschaften erfasst und sie grundlegend verändert. Auch vor den verschiedenen Disziplinen der Verwandtschafts- und Evolutionsforschung machte sie nicht halt. Bis dahin nutzten die Wissenschaftler vor allem Daten zum äußeren Aussehen, der Anatomie, der Zellen, der Embryonalentwicklung oder zur geographischen Verbreitung, um Verwandtschaftsverhältnisse (Systematik) zu untersuchen und Verwandtschaftslinien (Evolutionsgeschichte, Stammesgeschichte) zu rekonstruieren. Für viele dieser Merkmale war aber schon lange klar, dass sie dem verändernden Einfluss ihrer Umwelt (Selektion) unterworfen sind, was ihre evolutionäre und stammesgeschichtlich-systematische Interpretation erschwerte. Um dieses Problem zu lösen, wurden verschiedene wissenschaftliche Methoden entwickelt, die aber nicht zu einem zufriedenstellenden Ergebnis führten. Es gibt eigentlich nur zwei Auswege aus diesem Dilemma: Man analysiert ausschließlich Merkmale, die der Selektion nur wenig ausgesetzt sind. Oder man misst, wie sich die Selektion auf das betrachtete Merkmal auswirkt und macht deren Ausmaß quantifizierbar. Beides wird möglich, untersucht man das Erbmolekül, die DNS.
Die Gesamtheit der vererbbaren Informationen in der Zelle eines Individuums – das Erbgut oder „Genom“ – hat zwei wesentliche Eigenschaften, die es bei solchen Untersuchungen zu beachten gilt: Zum einen verändert sich das Genom über die Zeit durch Mutationen. Die Mutationsrate kann für bestimmte Bereiche des Genoms und über bestimmte Zeiträume hinweg konstant sein – in der Regel ist dies aber nicht der Fall. Zum zweiten darf das einzelne Genom nie isoliert betrachtet werden – es „ko-existiert“ vielmehr in den verschiedenen Individuen einer Population und Art, wird neu kombiniert und weitervererbt. Die Rekombination, das Durchmischen des Erbguts verschiedener Individuen bei sexuellen Vorgängen, ist dabei die erste wesentlich verändernde evolutionäre Größe – nicht die Mutationen. Und im Unterschied zur Mutation wird bei der Rekombination die Abfolge der Bausteine des Erbguts, die sogenannte DNS-Sequenz, primär nicht verändert.
Betrachtet man eine Population und nicht das einzelne Individuum liegt es nahe anzunehmen, dass auch die räumliche Verteilung der genetischen Variabilität dynamisch ist. In der Tat können alle Veränderungen des Erbguts benutzt werden, um die Geschichte der Evolution nachzuvollziehen. Das resultiert nicht nur in Stammbäumen von Arten, Gattungen und Familien; es führt auch zu einer völlig neuen Sichtweise der Evolutionsgeschichte einzelner Arten in „Zeit und Raum“: Genau das ist das Forschungsgebiet der Phylogeographie.
 |
|
|
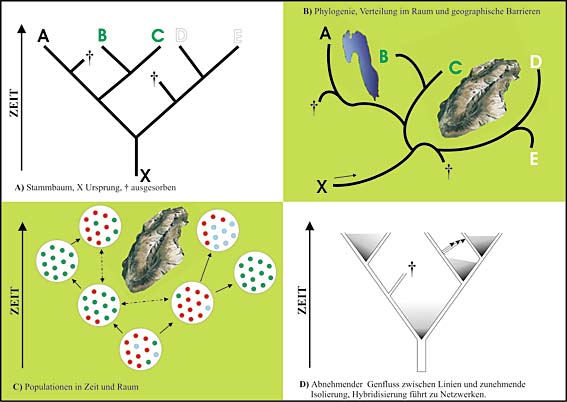
Häufig lassen sich Stammbäume von Arten (A) auf geographische Räume projizieren (B). Auch Populationen können solche Muster aufweisen (C), aber genetisch ist die Situation weitaus komplexer (D), da alle Übergänge von Genfluss bis zu völliger Isolation möglich sind.
|
Das ursprüngliche Grundprinzip phylogeographischer Rekonstruktion war, DNS-basierende Stammbäume oder Abstammungslinien mit der geographischen Verbreitung derjenigen taxonomischen Einheiten zu kombinieren, welche die zu betrachtenden DNS-Sequenzen tragen. Neue Konzepte lassen es nun zu, die phylogeographische Geschichte auch innerhalb einer Art zu rekonstruieren. Die zu untersuchende Einheit ist hier die Population, eine Gruppe von Individuen, die in engem Bezug steht (genetisch und/oder räumlich). Dabei spielt weniger die Mutation genetischer Information eine Rolle – entscheidend ist die Verteilung (Frequenz) einzelner DNS-Varianten (Allele) an ausgesuchten Genorten (Loci). Die Verteilung dieser Erbinformation ist ein Prozess, der sich in Zeit und Raum messen lässt (Populationsgenetik). Der Übergang der Phylogenetik, die sich mit Stammbäumen beschäftigt, zur Populationsgenetik, deren Augenmerk auf den Netzwerken liegt, ist dabei fließend.
Anhand von drei Beispielen – der Alpengänsekresse, des Hungerblümchens und des Gingko-Baumes – sollen einige Aspekte der neuen Forschungsrichtung vorgestellt werden. Es soll darüber hinaus gezeigt werden, wie es möglich wird, aktuelle genetische Daten zu nutzen, um in die Vergangenheit zu blicken und die Geschichte der Evolution zu rekonstruieren.
Die Alpengänsekresse Arabis alpina kommt, anders als es ihr Name vermuten lässt, nicht nur in den Alpen, sondern auch in der Tundra, in den Gebirgen Vorderasiens und in den Hochgebirgen Ostafrikas vor. Dieses Verbreitungsmuster scheint auf den ersten Blick einfach erklärbar: Arabis alpina überlebte die letzte Eiszeit im unvergletscherten Mitteleuropa und folgte dann den zurückweichenden Gletschern nach Süden in die alpinen Lebensräume und nach Norden bis in die Arktis. Es muss, so die Vorstellung, noch einen weiteren eiszeitlichen „Überlebensraum“ gegeben haben, der vom Balkan über das Ostmediterran bis nach Vorderasien reichte. Von dort gelangte die Art in einem bislang unbestimmt gebliebenen Zeitraum nach Afrika.
Soweit die „klassische“ Betrachtungsweise – die Kombination und Auswertung molekularer Daten zeigt uns indes ein völlig anderes Bild. Dazu muss man zunächst wissen, dass eine Pflanzenzelle über drei verschiedene Genome verfügt, die an unterschiedlichen Orten untergebracht sind: Das „Plastidengenom“ umfasst in höheren Landpflanzen etwa 150 000 Basenpaare (Erbgutbausteine), das „Mitochondriengenom“ hat eine in verschiedenen Pflanzenarten sehr variable Größe von 500 000 bis zwei Millionen Basenpaaren, das „Kerngenom“ besitzt mindestens 125 Millionen
Basenpaare.
Für die phylogeographische Betrachtung ist nun vor allem folgendes Charakteristikum bedeutsam: Die Plastiden werden bei bedecktsamigen Pflanzen (Angiospermen) in der Regel nur über die Mutterpflanzen vererbt. Da Pflanzen sich vorwiegend mit Samen ausbreiten – die von der Mutterpflanze gebildet werden –, beschreibt die Evolutionslinie der Plastiden nicht nur die „Plastiden-Evolution“, sondern auch die Ausbreitungsgeschichte der Pflanze im Raum. Wenn man mit molekularbiologischen Methoden die Abfolge der Basenpaare – die Sequenz – von Genen bestimmt, welche aus dem mütterlich vererbten Plastidengenom der Gänsekresse stammen, sind zahlreiche Varianten, sogenannte Haplotypen, zu erkennen, die in unterschiedlicher Art und Weise miteinander verwandt sind.
 |
|
|
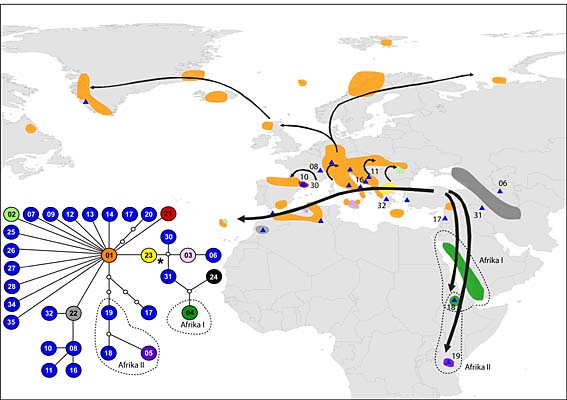
Die Verbreitung von Arabis alpina, der Alpengänsekresse, über den euroasiatischen und afrikanischen Raum lässt sich anhand der mütterlich vererbten Chloroplasten-DNS nachvollziehen. Die relative Verwandtschaft ist im Netzwerk aufgezeigt (im Bild links unten; * = vermutlicher evolutionärer Ursprung). Die kleinen Dreiecke repräsentieren Typen von Chloroplasten-DNS, die im gesamten Datensatz nur einmal aufgetreten sind.
|
Von diesen Daten lässt sich ablesen, dass die Alpengänsekresse ihren Ursprung nicht, wie ursprünglich angenommen, in Mitteleuropa, sondern in Kleinasien hat. Von dort hat sie ihre Wanderung vor mehr als 500.000 Jahren in Richtung Ostafrika angetreten. Diese Wanderrichtung hat sie während des Pleistozäns zweimal unabhängig voneinander eingeschlagen und sowohl die arabische Halbinsel wie Äthiopien besiedelt. Eine dritte Linie ist Richtung Mitteleuropa gewandert und hat dort alle Gebirge besiedelt. Die Eiszeiten haben die Art schließlich in südliche Refugien wie die Iberische Halbinsel, die Südwestalpen, die Apenninen oder den Balkan abgedrängt. Von dort folgte sie den rückweichenden Gletschern. DNS-Merkmale aus dem Kerngenom bestätigen dieses aus dem Plastidengenom ableitbare Szenario.
Wie und wann ist die Art aber nach Nordeuropa gelangt? Um diese Frage zu beantworten, haben wir gemeinsam mit der Arbeitsgruppe von Christian Brochmann in Oslo hochvariable DNS-Bereiche aus dem Kerngenom analysiert. Das Ergebnis war eine Überraschung: Während diejenigen Populationen, welche die Eiszeiten in den Berg- und Hügelgebieten Deutschlands überdauerten, genetisch unterschiedlich sind, haben alle Populationen in Skandinavien und im arktischen Verbreitungsgebiet den gleichen Genotyp. Diese genetische Einheitlichkeit lässt sich nur durch eine extrem schnelle, sehr effektive Ausbreitung einer kleinen Ausgangspopulation – womöglich gar eines einzelnen Individuums – kurz nach der letzten Eiszeit vor etwa 18000 Jahren erklären.
 |
|
|
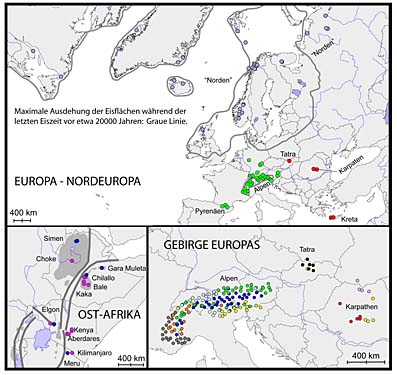
Die Grafik zeigt die genetische Zugehörigkeit von Populationen der Alpengänsekresse in unterschiedlichen Gebirgsregionen und anderen Großräumen. Die Daten dafür stammen aus der Analyse der im Kern enthaltenen Erbsubstanz.
|
Die genetische Verarmung hat die Alpengänsekresse offensichtlich nicht daran hindern können, in die nördlichen Lebensräume vorzudringen. Um solche Extremlebensräume in derart kurzer Zeit zu besiedeln, würde man eigentlich eine größere genetische Vielfalt als Voraussetzung erwarten. Doch auch populationsgenetische Berechnungen bestätigen die von uns ermittelten Zeiträume: Unter bestimmten Annahmen, etwa Populationsgröße und Generationsdauer, sowie der Zuhilfenahme von Mutationsraten ist die erfolgreiche Einwanderung der Pflanze von Zentralasien nach Mitteleuropa mit einiger Wahrscheinlichkeit in die letzte Zwischeneiszeit, das „Eem Interglazial“ (vor etwa 126 000 bis 115 000 Jahren), zu datieren. Die Besiedlung Afrikas von Kleinasien aus geschah vor etwa 500 000 Jahren und fällt damit nicht in das Tertiär. Damit ist auch die klassische These, nach der die alpine Vegetation der ostafrikanischen Gebirge eine Tertiärflora ist, generell in Frage gestellt.
Die Phylogeographie der Alpengänsekresse zeigt uns, dass eine hohe genetische Variabilität über lange Zeit grundlegend sein kann für eine geographisch weite und langfristig erfolgreiche Besiedlung in einer sich verändernden Umwelt. Dennoch kann eine Art in vergleichsweise kurzen Zeiträumen von wenigen Zehntausenden von Jahren trotz genetisch geringer Variabilität äußerst erfolgreich sein. Es wird spannend sein zu erfahren, ob und wie zukünftige gravierende Umweltveränderungen die Art im äußersten Norden beeinflussen werden. Möglicherweise helfen unsere Erkenntnisse besser zu verstehen, wie es Pflanzen gelingt, sich an einen schnellen Klimawandel anzupassen.
 |
|
|
„Frucht“ des Gingko-Baumes, aufgenommen im Botanischen Garten der Universität Heidelberg
|
rekonstruieren. In der frühen Evolution der Gruppe entstehen vor etwa fünf Millionen Jahren verschiedene Linien, die sehr unterschiedliche Lebensräume besiedeln und eine Reihe einjähriger Arten umfassen. Mit der ersten dramatischen Klimaveränderung zu Beginn des Pleistozäns vor zwei Millionen Jahren bildet sich dann der erste große Artengrundstock der mehrjährigen alpinen und arktischen Extremisten in einem bemerkenswerten Artbildungsprozess, einer sogenannten Radiation. Während dieses Prozesses ist die Artbildungsrate – also das Entstehen neuer Arten abzüglich der in dieser Zeiteinheit aussterbenden Arten – plötzlich und drastisch erhöht. Eine dritte Radiationswelle lässt sich für die Mitte des Pleistozäns berechnen. Bemerkenswert ist, dass die dramatischen Klimaveränderungen bei dieser Gattung die höchsten heute im Pflanzenreich bekannten Artbildungsraten ausgelöst haben.
Was unterscheidet nun das Modell „Draba“ vom Modell „Arabis alpina“? Im Falle der Hungerblümchen betrachten wir größere Zeiträume, die Arten sind um den Erdball gewandert, sie haben sich dabei genetisch verändert und Mutation und Selektion haben in einer sich verändernden Umwelt neue Arten hervorgebracht, die den klimatischen Schwankungen mit nachfolgenden großen Arealverschiebungen „hinterherlaufen“ mussten. Wie lässt sich das erklären? Wo kommt das gewaltige genetische Potenzial her, das diese ständig neuen Anpassungen ermöglicht?
Im Falle der Gattung Draba lauten die Antworten auf diese Fragen „Hybridisierung“ und „Polyploidisierung“. Mit Hybridisierung ist die genetische Kreuzung verschiedener Arten gemeint; der Fachbegriff Polyploidisierung bezeichnet das Vervielfachen des gesamten Genoms in jeder Zelle: Damit gelingt es Pflanzen, verschiedene Erbgutsätze, die durch Hybridisierung zusammengekommen sind, genetisch zu stabilisieren. Wir können zeigen, dass die vielen Draba-Arten gleichsam als große Gruppen über die Nordhemisphäre gewandert sind, wobei sie Gletschern weichen mussten und immer wieder aus vereisten Gebirgen herausgedrängt wurden. Die Arten folgten aber auch den sehr rasch zurückweichenden Gletschern, und zwar sowohl in die Höhe als auch in Richtung Norden. Diese Dynamik bedingte, dass verschiedene Arten immer wieder miteinander in Kontakt kamen, hybridisierten und ihr neu kombiniertes Erbgut mithilfe der Polyploidisierung stabilisierten. Tatsächlich sehen wir bei der Gattung Draba eine starke Korrelation von Ploidiegrad und Höhenverbreitung beziehungsweise Verbreitung in Richtung Arktis.
Die Gattung Draba ist sicherlich ein evolutionärer Sonderfall, denn sie hat während ihrer nordhemisphärischen Wanderungen zudem „evolutionäre Ringe“ gebildet: Es gibt Wanderungslinien, die sich von Vorder- und Zentralasien über den Himalaja weiter Richtung Beringstraße und den nordamerikanischen Kontinent ausgebreitet haben. Dort sind die Ausbreitungslinien mehrfach auf eine evolutionäre Linie getroffen, die von Europa nach Norden, in die Arktis, Richtung Grönland und Nordostamerika gewandert ist. Die dabei entstandenen neuen („hybridogenen“) Evolutionslinien sind dann mehrmals unabhängig voneinander die Anden entlang nach Süden bis zu den Falklandinseln gewandert. Ähnliche, wenn auch kleinere „Kreise“ finden sich beispielsweise um die Gebirge in Zentral- und Ostasien. Vordergründig scheint die Gattung Draba also ein Beispiel dafür zu sein, wie dramatische klimatische Veränderungen die Bildung neuer biologischer Vielfalt fördern und die Vielfalt des Lebens insgesamt erhöhen. Genauer betrachtet sind es aber nicht die Veränderungen der Umwelt, die dies bewirken, sondern der ständige Wechsel und die Möglichkeit, auf den Wandel zeitlich und räumlich zu „reagieren“.
 |
|
|
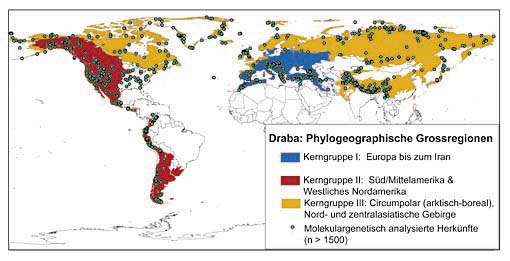
Die Verbreitung des Hungerblümchens (Gattung Draba). Mit den Farben blau, rot und gelb unterlegt ist die stammesgeschichtliche Zuordnung zu einer der drei evolutionären Hauptlinien.
|
Über die Evolutionsgeschichte des Gingko ist wenig bekannt. Es ist vor allem eine Geschichte, die von Fossilfunden erzählt wird. Der Gingko ist ein „lebendes Fossil“, der einzige Vertreter einer evolutionär völlig isoliert stehenden Entwicklungslinie unter den noch heute lebenden Samenpflanzen. Gingko-ähnliche Bäume lassen sich bis in das frühe Perm vor etwa 280 Millionen Jahre zurückverfolgen. Die Gattung Gingko erscheint dann im frühen Jura vor etwa 180 Millionen Jahren und erreicht ihre größte Artenvielfalt im späten Mesozoikum und frühen Tertiär vor etwa 120 Millionen Jahren mit einer weiten Verbreitung auf der gesamten Nordhemisphäre. Vor etwa sieben bis zehn Millionen Jahren verschwand der Gingko in Nordamerika, die letzten in Europa gefundenen Fossilien sind 1,7 bis 2,7 Millionen Jahre alt.
Nach den Fossilienfunden zu urteilen, starb der Gingko in Europa zu Beginn des Eiszeitalters aus. Alle weiteren Fossilien aus dem späten Tertiär und dem frühen Pleistozän stammen aus Ostasien. Nach diesem sich über den gesamten Erdball fortsetzenden Aussterben trat der Gingko als Gingko biloba in Mitteleuropa erst wieder vor etwa 250 Jahren auf: Engelbert Kämpfer, ein deutscher Arzt und Botaniker, brachte den Gingko im Jahr 1692 aus Japan als Samen mit, der erste Gingko-Baum wurde im Botanischen Garten von Leiden gepflanzt. Bis dahin galt der Gingko zumindest in der westlichen Welt als ausgestorben. In China, Taiwan, Korea und Japan aber war der Gingko seit Jahrtausenden ein heiliger Baum und wurde durch den Menschen weit verbreitet.
Lange war gänzlich unbekannt, in welchen Arealen der Gingko überlebt haben sollte. Die Analyse der Erbinformation der Plastiden- und der Kern-DNS lässt nun auf zwei eiszeitliche Überlebensareale rückschließen. Und zwar auf ein großes Gebiet mit ursprünglichen gemäßigt-submediterranen Laubwäldern im Südwesten Chinas und ein kleineres Areal in Ostchina (Mt. Tianmu, Zhejiang). Die Analyse der genetischen Struktur der Populationen hat ergeben, dass von diesen Überlebensarealen ausgehend der Gingko seinen „Eroberungszug“ – vornehmlich mithilfe des Menschen – in wenig mehr als 2000 Jahren über China, Korea und Japan und schließlich weiter über Europa und Amerika vollzogen hat.
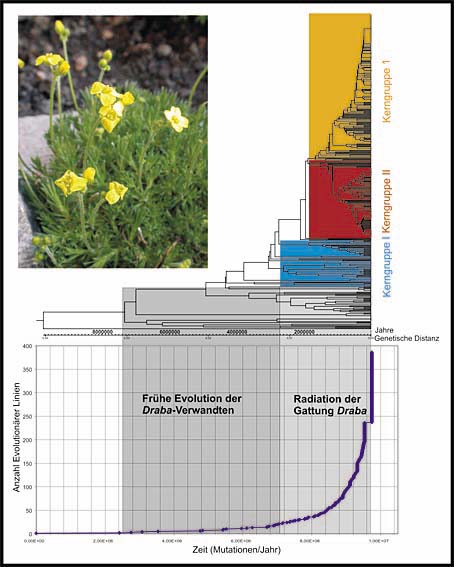 |
|
|
Der Stammbaum der Gattung Draba, auf ein Diagramm übertragen: Es zeigt die evolutionären Hauptlinien zu einem bestimmten Zeitpunkt in der Vergangenheit; je steiler der Graph ausfällt, desto schneller verläuft die Artbildung.
|
Die Geschichte des Gingko ist deshalb so bemerkenswert, weil der Baum, der ein Alter von mehr als tausend Jahren erreichen kann, als besonders „industriefest“ gilt und ein beliebter Stadtbaum ist. Man würde ihm auf den ersten Blick also eine besonders große Überlebensfähigkeit zubilligen wollen. Die Evolutionsgeschichte aber zeigt uns ganz im Gegenteil, dass der Gingko kurz davor steht, endgültig auszusterben – sofern er vom Menschen nicht weiterhin kultiviert und gefördert wird.
Die Evolutionsgeschichten von der Alpengänsekresse, vom Hungerblümchen und vom Gingko-Baum demonstrieren exemplarisch, dass in Zeiten großräumiger Umweltveränderungen einige wenige Arten und Artengruppen gefördert werden und die biologische Viel-falt hier partiell steigt, viele andere Organismen aber aussterben oder nur wenige Vertreter in bestimmten Arealen überleben. Die Analyse genetischer Veränderungen, die in der Vergangenheit der Arten und Artengruppen stattgefunden haben, erlaubt es uns erstmals, evolutionäre Prozesse konkret zu rekonstruieren und zu erkennen, welche Merkmale und Anpassungen für den Erfolg oder Misserfolg von Organismen im Wandel der Zeiten verantwortlich waren. Solche, anhand vergangener Ereignisse erkannte Schlüsselmerkmale und Schlüsseleigenschaften können alsdann in heute noch lebenden Arten der entsprechenden Verwandtschaftsgruppe untersucht werden. Auf diese Weise lässt der Blick zurück gleichzeitig in die Zukunft sehen – und die Frage beantworten, wie Pflanzen auf Veränderungen, beispielsweise den derzeitigen Klimawandel, reagieren und ob sie ihn überstehen werden.
 |
|
Kontakt: mkoch@hip.uni-heidelberg.de